Las bacterias son los organismos más pequeños que poseen toda la maquinaria requerida para su crecimiento y replicación, son procariotas y presentan formas celulares características.
Sor organismos relativamente simples y algunos son fáciles de manipular en el laboratorio, son importantes en la ecología de la tierra ya que participan en la fijación del nitrógeno, son eje central del ciclo del carbono debido a su habilidad para degradar muchos polímeros naturales como la celulosa y otros compuestos.
Morfología:
Principales grupos morfológicos:
Cocos: forma esférica u ovoide, pueden existir como células individuales o en agrupaciones:
Sor organismos relativamente simples y algunos son fáciles de manipular en el laboratorio, son importantes en la ecología de la tierra ya que participan en la fijación del nitrógeno, son eje central del ciclo del carbono debido a su habilidad para degradar muchos polímeros naturales como la celulosa y otros compuestos.
Morfología:
Principales grupos morfológicos:
Cocos: forma esférica u ovoide, pueden existir como células individuales o en agrupaciones:

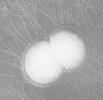
Diplococos: se forman cuando dos cocos se dividen y permanecen juntos formando pares. (Neisseria sp.)
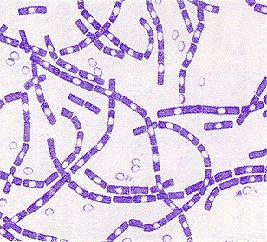
Cadenas: formación de largas filas (Streptococcus sp.)
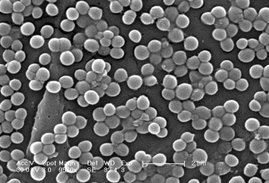
Racimos: agrupaciones en forma de racimos de uva (Staphylococcus sp.)
Bacilos: forma cilíndrica, varían en longitud y anchura, el extremo puede ser plano, redondeado o bifurcado. Pueden formar parejas o cadenas. Algunos pueden ser curvos.
Cadenas: formación de largas filas (Streptococcus sp.)
Racimos: agrupaciones en forma de racimos de uva (Staphylococcus sp.)
Bacilos: forma cilíndrica, varían en longitud y anchura, el extremo puede ser plano, redondeado o bifurcado. Pueden formar parejas o cadenas. Algunos pueden ser curvos.

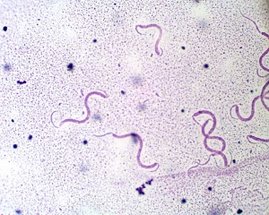
Espirilios: bacilos que se curvan en forma espiral

Espiroquetas: en forma de sacacorchos

Filamentosas: células largas y delgadas, o cadenas de células.


Los procariotas presentan tamaños que van desde 0.1-0.2 μm de ancho y 50 μm de diámetro. Para efectos comparativos, una célula eucariota típica puede variar de 2 a más de 200 μm de diámetro, por lo tanto la mayoría de los procariotas son muchos más pequeños que los eucariotas y el tamaño pequeño determina algunas de sus propiedades biológicas.
Por ejemplo la velocidad con la que los nutrientes penetran al interior de la célula y salen de esta las sustancias de desecho constituye un factor clave porque influye en los ritmos metabólicos celulares y en la velocidad de crecimiento. La relación es inversamente proporcional al tamaño celular.
Las velocidades de transporte son en parte dependientes de la superficie de membrana disponible respecto al volumen. Con relación a su tamaño, las células pequeñas tienen más superficie relativa disponible que las grandes, por lo que puede llevar a cabo un intercambio de nutrientes con el medio en condiciones más ventajosas.
Las células pequeñas manifiestan velocidades de crecimiento más rápidas y la formación de poblaciones celulares más grandes respecto a las células de mayor tamaño.
Estructuras internas:

Por ejemplo la velocidad con la que los nutrientes penetran al interior de la célula y salen de esta las sustancias de desecho constituye un factor clave porque influye en los ritmos metabólicos celulares y en la velocidad de crecimiento. La relación es inversamente proporcional al tamaño celular.
Las velocidades de transporte son en parte dependientes de la superficie de membrana disponible respecto al volumen. Con relación a su tamaño, las células pequeñas tienen más superficie relativa disponible que las grandes, por lo que puede llevar a cabo un intercambio de nutrientes con el medio en condiciones más ventajosas.
Las células pequeñas manifiestan velocidades de crecimiento más rápidas y la formación de poblaciones celulares más grandes respecto a las células de mayor tamaño.
Estructuras internas:

Membrana citoplasmática:
Se localiza alrededor del citoplasma por debajo de la pared celular. Constituye una barrera diferencial selectivamente permeable que separa el lado interno del externo, por lo que regula el transporte de materiales hacia adentro o hacia afuera de la célula.
Está constituida por:
- Fosfolípidos 20-30%
- Proteínas 60-70%
Las moléculas de fosfolípidos tienen terminaciones polares (hidrofílicas) y terminaciones no polares (hidrofóbicos) por lo que se denominan antipáticos. Se organizan de una manera tal que forman una bicapa fosfolipídica con sus extremos polares hacia el exterior y los no polares hacia el interior.
Esta bicapa provee una barrera hidrofóbica que es solamente permeable al agua y a pequeñas moléculas hidrofóbicas.
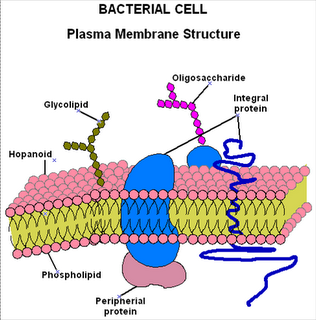
En las Eubacterias los fosfolípidos son fosfoglicéridos en donde las cadenas de ácidos grasos se unen al glicerol por enlaces tipo éster (-CO-O), en las Arquibacterias existen lípidos compuestos por dos cadenas de hidrocarburos unidos por un enlace éter (-O-) con el glicerol, lo que conforma un diéter compuesto de glicerol y dos moléculas de alcohol (fitanol).
Las membranas bacterianas se diferencian normalmente de las eucariotas en que carecen de esteroles como colesterol (a excepción de los Mycoplasma). Sin embargo, muchas membranas contienen moléculas pentacíclicas, similares al esterol, denominadas hopanoides. Estas sustancias, del mismo modo que los esteroides en los eucariotas, probablemente estabilicen la membrana bacteriana.
El modelo más aceptado actualmente es el modelo de Mosaico Fluido, que puede presentar cambios reversibles de estado dependiendo de la temperatura y la composición.
Existen dos tipos de proteínas:
- Integrales 70-80%
- Periféricas 20-30%
Las proteínas integrales están unidas firmemente a la membrana por lo que, no se extraen fácilmente. Las proteínas periféricas se encuentran asociadas a la superficie y pueden se removidas por choque osmótico. El tipo de proteínas presentes en la membrana varían de acuerdo a las condiciones de crecimiento del organismo (aerobias o anaerobias).
La principal función de la membrana citoplasmática es la de regular el flujo de materiales dentro y fuera de la célula. Los mecanismos por los cuales se llevan a cabo estos procesos son la difusión pasiva, el transporte activo, la translocación de grupos y la citosis.
La membrana plasmática bacteriana es también el lugar donde se desarrollan numerosos procesos metabólicos: respiración, fotosíntesis, síntesis de lípidos y de constituyentes de la pared celular.
Mesosomas:
Invaginaciones de la membrana plasmática para formar vesículas, túmulos o lamelas y se observan tanto en bacterias Gram negativas como en Gram positivas, siendo más prominentes en estas últimas.
Se desconoce su función exacta, pero como suelen situarse a continuación de los septos o tabiques que dividen las bacterias y a veces parecen estar unidas al cromosoma bacteriano se cree que pueden estar involucrados en la formación de la pared celular durante la división o desarrollar un papel en la replicación del cromosoma y su distribución en las células hijas.
También, pueden estar involucrados en procesos secretorios.
Citoplasma:
A diferencia del citoplasma eucariota, este carece de organelas provistas de membranas.
Constituido principalmente por agua (por lo menos el 70% de la masa bacteriana es agua), tiene la apariencia semejante a un gel y se encuentra englobada en la membrana citoplasmática.
Su función es la de retener componentes celulares donde se llevan a cabo muchas de las funciones de crecimiento, metabolismo y replicación.
Existen regiones de interés como:
- Región cromatídica que es rica en ADN
- Región granular formada por los ribosomas
- Región fluida donde están disueltas las sustancias.
Cuerpos de Inclusión:
Son gránulos de material orgánico o inorgánico en el citoplasma que son usualmente visibles al microscopio. Poseen una composición variada algunas de naturaleza proteínica y otras pueden contener lípidos. Su importancia es que en condiciones favorables sirve de almacenaje de sustancias que posteriormente le sirven como reservas nutritivas.
Cuerpos de inclusión orgánicos:
- Glucógeno: polímero de unidades de glucosa Reserva de carbono
- Poli-β-hidroxibutirato: moléculas de β-hidroxibutirato para energía y biosíntesis
- Gránulos de cianomicina: producidas por las cianobacterias, compuestos de grandes polipéptidos con partes iguales arginina y ácido aspártico. Almacenan nitrógeno extra para la bacteria.
- Carboxisomas: cianobacterias, bacterias nitrificantes y tiobacilos. Son inclusiones poliédricas que contienen la enzima ribulosa-1,5-bifosfato carboxilasa, y sirve como reserva de la misma o como sitio para la fijación de CO2.
- Vacuolas de gas: cianobacterias, otras bacterias fotosintéticas y formas acuáticas (Holobacterium). Estas bacterias pueden flotar en o cerca de la superficie porque estas vacuolas les da flotabilidad.
Cuerpos de inclusión inorgánicos:
- Gránulos de polifosfato: Polímero lineal de ortofosfatos que sirven como fuente de energía.
- Gránulos de Volutina: reserva de fosfato.
Ribosomas:
Los ribosomas son pequeños y aunque parezcan partículas sin ninguna característica importante son muy complejos. De composición proteica (35%) y de ácido ribonucleico (65%) son el sito de síntesis de proteínas.
Son más pequeños que los ribosomas eucariotas y se denominan comúnmente ribosomas 70S (subunidades 50S y 30S).
Nucleoide:
El material genético no se encuentra delimitado por una membrana nuclear, el cromosoma procariótico (ADN doble banda circular) está localizado en una región de forma irregular llamada nucleoide.
Análisis químicos han revelado que está compuesto de 60% de ADN, algo de ARN y una pequeña cantidad de proteína.
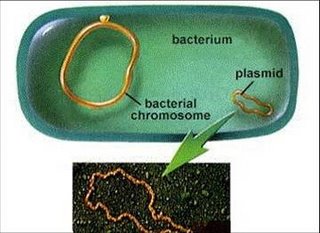
Plámidos:
Las bacterias pueden contener, además del cromosoma, una o más moléculas circulares de ADN doble banda, llamada plásmidos que contienen una cantidad limitada de información genética específica.
Pueden replicarse independientemente o pueden estar integrados en el cromosoma. No son necesarios para el crecimiento o reproducción de la bacteria, pero pueden aportar genes que aportan una ventaja selectiva (resistencia a antibióticos, nuevas habilidades metabólicas, patogenicidad).
Endosporas:
Estructuras únicas de las bacterias (principalmente gram positivas), los principales géneros: Bacillus, Clostridium, Sporosarcina, Desulfotomoculum y Thermoactinomyces.
Son cuerpos refráctiles que no se tiñen con los colorantes ordinarios y son resistentes al calor, a la desecación, radiación, agentes desinfectantes y otros agentes ambientales que puedan afectar el crecimiento bacteriano. Representan un avance selectivo para la supervivencia y la diseminación de las especies que son capaces de originarlas.
La forma y localización depende de la especie bacteriana, por forma pueden ser ovoides, esféricas o elipsoidales y por su localización pueden ser terminales, centrales o subterminales.
La esporulación se produce al cesar el crecimiento celular como consecuencia del gasto de nutrientes, existen diversos cambios en la expresión de genes de la célula (Bacillus al menos 200 genes están implicados en el proceso). Requiere el cese de la síntesis de algunas proteínas implicadas en las funciones celulares y la producción de proteínas específicas de la espora.

Rodeada por una capa delicada y delgada de naturaleza proteica denominada exosporio. Debajo de este se encuentra la cubierta de la espora compuesta por varias capas de proteínas. Bajo la cubierta descansa el córtex constituido por un peptidoglicano modificado (menor número de enlaces). Debajo la pared que rodea al protoplasto que contiene las estructuras celulares nomales.
Las capas contienen mureína y dipicolinato de calcio (15% del peso seco) que es el responsable de la resistencia al calor al estabilizar los ácidos nucleicos.
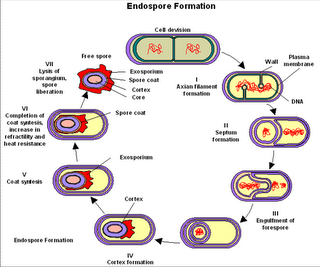
Esporulación:
- Fase 0: se duplica el material genético
- Fase I: formación del filamento axial
- Fase II: plegamiento interno de la membrana celular para englobar uno de los cromosomas y producir el septo de la preespora
- Fase III: membrana continúa creciendo y engloba la espora inmadura con una segunda membrana.
- Fase IV: Se forma el córtex entra las 2 membranas, donde se acumula el dipicolinato de calcio.
- Fase V: se forman las cubiertas
- Fase VI: maduración de la espora
- Fase VII: enzimas líticas liberan la espora
Paso de espora a célula vegetativa:
Activación: proceso reversible, preparación de la espora para la germinación, se da por algún estímulo, por ejemplo el calor.
Germinación: es un proceso rápido, se hincha, la cubierta se rompe o se absorbe. Se pierde la resistencia al calor, radiación, desecación, etc. Se desencadena por ciertos metabolitos, por ejemplo los nutrientes del medio (aminoácidos, azúcares, etc.).
Crecimiento: protoplasto fabrica nuevos componentes.
Estructuras externas:
Pared celular:
Las bacterias, a excepción de algunos micoplasmas y algunas arquibacterias, tienen una pares fuerte que les da forma y las protege de la lisis osmótica y sustancias tóxicas. En algunos microorganismos puede tener componentes que le confieren patogenicidad.
En 1884 el investigador danés Christian Gram observó diferencias tintoriales relacionadas con la habilidad de ciertas células bacterianas de retener el colorante cristal violeta después de la decoloración con etanol al 95%, estas células fueron llamadas Gram positivas. Las células que se decoloraban y solamente son visibles al teñirlas con un colorante de contraste son referidas como Gram negativas.
Esto se debía a las diferencias existentes en la estructura de la pared celular:
- Gram positiva: formada por una única capa homogénea de 20-80 nm de grosor, de peptidoglucano o mureína situado por fuera de la membrana plasmática. Además de polímeros aniónicos (ácidos teicoicos y teicourónicos), polisacáridos neutros, proteínas y en algunos casos lípidos. Los ácidos teicoicos se encuentran presentes en la pared de dos maneras, unidas covalentemente al peptidoglucano o a lípidos de membrana como ácidos lipoteicoicos. Sus funciones son unir la pared celular con la membrana citoplasmática y acumular cargan negativas para atrapar cationes como el Mg2+, que es esencial para la actividad normal de las enzimas de membrana. Existe un espacio periplásmico que no tiene tantas proteínas como en las Gram negativas, pero secretan exoenzimas.

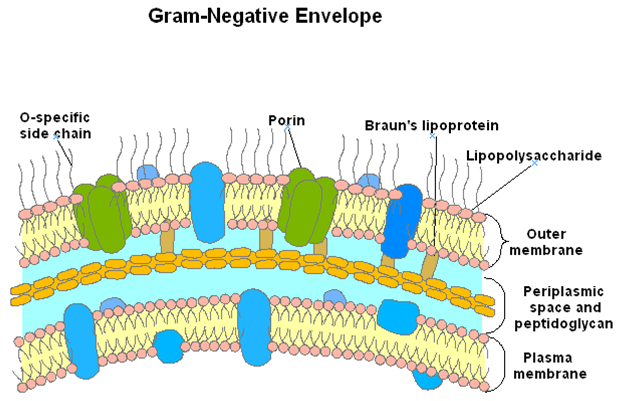
- Gram negativa: es más compleja, posee una capa de 2-7 nm de grosor de peptidoglucano rodeada por una membrana externa de unos 7-8 nm. El espacio entre estas dos capas se denomina espacio periplásmico y la sustancia que ocupa ese lugar periplasma. En este espacio hay muchas proteínas que participan en la captación de nutrientes y proteínas quelantes que participan en el transporte de materiales hacia el interior de la célula, contiene también enzimas que participan en la síntesis del peptidoglucano y en la modificación de compuestos tóxicos que podrían lesionar a la célula.
Por fuera de la fina capa de peptidoglucano se encuentra una membrana externa que cumple la función de una membrana lisosomal, las enzimas hidrolíticas del espacio periplásmico se mantienen separadas de la célula gracias a la membrana y al peptidoglucano para prevenir la autodigestión.
El espacio periplásmico es isoosmótico con el citoplasma y fuertemente aniónico con el medio externo, por lo que es en parte responsable de mantener la osmolaridad (afuera es hiposomótico). La membrana externa funciona como una barrera permeable para los solutos hidrofilicos.
El componente más importante de la membrana externa es el Lipopolisacárido (LPS) o endotoxina que es la principal toxina de las bacterias entéricas patógenas. Los fosfolípidos y proteínas se encuentran en menor concentración que en la membrana plasmática. Existe un tipo de proteínas llamadas porinas que forman canales transmembrana y permiten el paso de solutos de acuerdo al peso molecular (nutrientes hidrofílicos).

Peptidoglucano: Polímero constituido por residuos alternantes de N-acetilglucosamina y ácido N- acetilmurámico (aminoazúcares). Una cadena de 4 aminoácidos D- y L- alternantes está conectada al ácido N- acetilmurámico y varía de acuerdo a las especies.

Glicocálix:
Polisacáridos de origen bacteriano que se localizan alrededor de la membrana externa (Gram negativas) y del peptidoglucano Gram positivas).
Se clasifican como:
- Slime o tipo S: matriz difusa
- Cápsula: matriz fibrosa bien organizada:
rígida
flexible
integral
periférica
La cápsula está formada por polisacáridos y en algunos casos puede contener proteínas, su componente principal es el agua lo que puede proteger a la bacteria de la desecación. Permite a las bacterias unirse a diferentes tipos de sustratos, previene la ingestión de la bacteria por las células fagocíticas, evita virus bacterianos y puede dar patogenicidad.

Se localiza alrededor del citoplasma por debajo de la pared celular. Constituye una barrera diferencial selectivamente permeable que separa el lado interno del externo, por lo que regula el transporte de materiales hacia adentro o hacia afuera de la célula.
Está constituida por:
- Fosfolípidos 20-30%
- Proteínas 60-70%
Las moléculas de fosfolípidos tienen terminaciones polares (hidrofílicas) y terminaciones no polares (hidrofóbicos) por lo que se denominan antipáticos. Se organizan de una manera tal que forman una bicapa fosfolipídica con sus extremos polares hacia el exterior y los no polares hacia el interior.
Esta bicapa provee una barrera hidrofóbica que es solamente permeable al agua y a pequeñas moléculas hidrofóbicas.
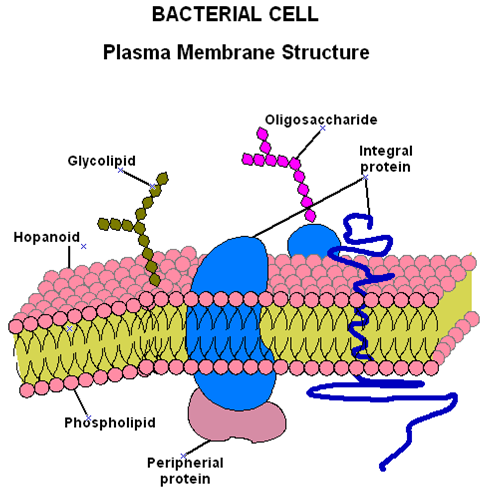
En las Eubacterias los fosfolípidos son fosfoglicéridos en donde las cadenas de ácidos grasos se unen al glicerol por enlaces tipo éster (-CO-O), en las Arquibacterias existen lípidos compuestos por dos cadenas de hidrocarburos unidos por un enlace éter (-O-) con el glicerol, lo que conforma un diéter compuesto de glicerol y dos moléculas de alcohol (fitanol).
Las membranas bacterianas se diferencian normalmente de las eucariotas en que carecen de esteroles como colesterol (a excepción de los Mycoplasma). Sin embargo, muchas membranas contienen moléculas pentacíclicas, similares al esterol, denominadas hopanoides. Estas sustancias, del mismo modo que los esteroides en los eucariotas, probablemente estabilicen la membrana bacteriana.
El modelo más aceptado actualmente es el modelo de Mosaico Fluido, que puede presentar cambios reversibles de estado dependiendo de la temperatura y la composición.
Existen dos tipos de proteínas:
- Integrales 70-80%
- Periféricas 20-30%
Las proteínas integrales están unidas firmemente a la membrana por lo que, no se extraen fácilmente. Las proteínas periféricas se encuentran asociadas a la superficie y pueden se removidas por choque osmótico. El tipo de proteínas presentes en la membrana varían de acuerdo a las condiciones de crecimiento del organismo (aerobias o anaerobias).
La principal función de la membrana citoplasmática es la de regular el flujo de materiales dentro y fuera de la célula. Los mecanismos por los cuales se llevan a cabo estos procesos son la difusión pasiva, el transporte activo, la translocación de grupos y la citosis.
La membrana plasmática bacteriana es también el lugar donde se desarrollan numerosos procesos metabólicos: respiración, fotosíntesis, síntesis de lípidos y de constituyentes de la pared celular.
Mesosomas:
Invaginaciones de la membrana plasmática para formar vesículas, túmulos o lamelas y se observan tanto en bacterias Gram negativas como en Gram positivas, siendo más prominentes en estas últimas.
Se desconoce su función exacta, pero como suelen situarse a continuación de los septos o tabiques que dividen las bacterias y a veces parecen estar unidas al cromosoma bacteriano se cree que pueden estar involucrados en la formación de la pared celular durante la división o desarrollar un papel en la replicación del cromosoma y su distribución en las células hijas.
También, pueden estar involucrados en procesos secretorios.
Citoplasma:
A diferencia del citoplasma eucariota, este carece de organelas provistas de membranas.
Constituido principalmente por agua (por lo menos el 70% de la masa bacteriana es agua), tiene la apariencia semejante a un gel y se encuentra englobada en la membrana citoplasmática.
Su función es la de retener componentes celulares donde se llevan a cabo muchas de las funciones de crecimiento, metabolismo y replicación.
Existen regiones de interés como:
- Región cromatídica que es rica en ADN
- Región granular formada por los ribosomas
- Región fluida donde están disueltas las sustancias.
Cuerpos de Inclusión:
Son gránulos de material orgánico o inorgánico en el citoplasma que son usualmente visibles al microscopio. Poseen una composición variada algunas de naturaleza proteínica y otras pueden contener lípidos. Su importancia es que en condiciones favorables sirve de almacenaje de sustancias que posteriormente le sirven como reservas nutritivas.
Cuerpos de inclusión orgánicos:
- Glucógeno: polímero de unidades de glucosa Reserva de carbono
- Poli-β-hidroxibutirato: moléculas de β-hidroxibutirato para energía y biosíntesis
- Gránulos de cianomicina: producidas por las cianobacterias, compuestos de grandes polipéptidos con partes iguales arginina y ácido aspártico. Almacenan nitrógeno extra para la bacteria.
- Carboxisomas: cianobacterias, bacterias nitrificantes y tiobacilos. Son inclusiones poliédricas que contienen la enzima ribulosa-1,5-bifosfato carboxilasa, y sirve como reserva de la misma o como sitio para la fijación de CO2.
- Vacuolas de gas: cianobacterias, otras bacterias fotosintéticas y formas acuáticas (Holobacterium). Estas bacterias pueden flotar en o cerca de la superficie porque estas vacuolas les da flotabilidad.
Cuerpos de inclusión inorgánicos:
- Gránulos de polifosfato: Polímero lineal de ortofosfatos que sirven como fuente de energía.
- Gránulos de Volutina: reserva de fosfato.
Ribosomas:
Los ribosomas son pequeños y aunque parezcan partículas sin ninguna característica importante son muy complejos. De composición proteica (35%) y de ácido ribonucleico (65%) son el sito de síntesis de proteínas.
Son más pequeños que los ribosomas eucariotas y se denominan comúnmente ribosomas 70S (subunidades 50S y 30S).
Nucleoide:
El material genético no se encuentra delimitado por una membrana nuclear, el cromosoma procariótico (ADN doble banda circular) está localizado en una región de forma irregular llamada nucleoide.
Análisis químicos han revelado que está compuesto de 60% de ADN, algo de ARN y una pequeña cantidad de proteína.

Plámidos:
Las bacterias pueden contener, además del cromosoma, una o más moléculas circulares de ADN doble banda, llamada plásmidos que contienen una cantidad limitada de información genética específica.
Pueden replicarse independientemente o pueden estar integrados en el cromosoma. No son necesarios para el crecimiento o reproducción de la bacteria, pero pueden aportar genes que aportan una ventaja selectiva (resistencia a antibióticos, nuevas habilidades metabólicas, patogenicidad).
Endosporas:

Estructuras únicas de las bacterias (principalmente gram positivas), los principales géneros: Bacillus, Clostridium, Sporosarcina, Desulfotomoculum y Thermoactinomyces.
Son cuerpos refráctiles que no se tiñen con los colorantes ordinarios y son resistentes al calor, a la desecación, radiación, agentes desinfectantes y otros agentes ambientales que puedan afectar el crecimiento bacteriano. Representan un avance selectivo para la supervivencia y la diseminación de las especies que son capaces de originarlas.
La forma y localización depende de la especie bacteriana, por forma pueden ser ovoides, esféricas o elipsoidales y por su localización pueden ser terminales, centrales o subterminales.
La esporulación se produce al cesar el crecimiento celular como consecuencia del gasto de nutrientes, existen diversos cambios en la expresión de genes de la célula (Bacillus al menos 200 genes están implicados en el proceso). Requiere el cese de la síntesis de algunas proteínas implicadas en las funciones celulares y la producción de proteínas específicas de la espora.
Rodeada por una capa delicada y delgada de naturaleza proteica denominada exosporio. Debajo de este se encuentra la cubierta de la espora compuesta por varias capas de proteínas. Bajo la cubierta descansa el córtex constituido por un peptidoglicano modificado (menor número de enlaces). Debajo la pared que rodea al protoplasto que contiene las estructuras celulares nomales.
Las capas contienen mureína y dipicolinato de calcio (15% del peso seco) que es el responsable de la resistencia al calor al estabilizar los ácidos nucleicos.
Esporulación:
- Fase 0: se duplica el material genético
- Fase I: formación del filamento axial
- Fase II: plegamiento interno de la membrana celular para englobar uno de los cromosomas y producir el septo de la preespora
- Fase III: membrana continúa creciendo y engloba la espora inmadura con una segunda membrana.
- Fase IV: Se forma el córtex entra las 2 membranas, donde se acumula el dipicolinato de calcio.
- Fase V: se forman las cubiertas
- Fase VI: maduración de la espora
- Fase VII: enzimas líticas liberan la espora
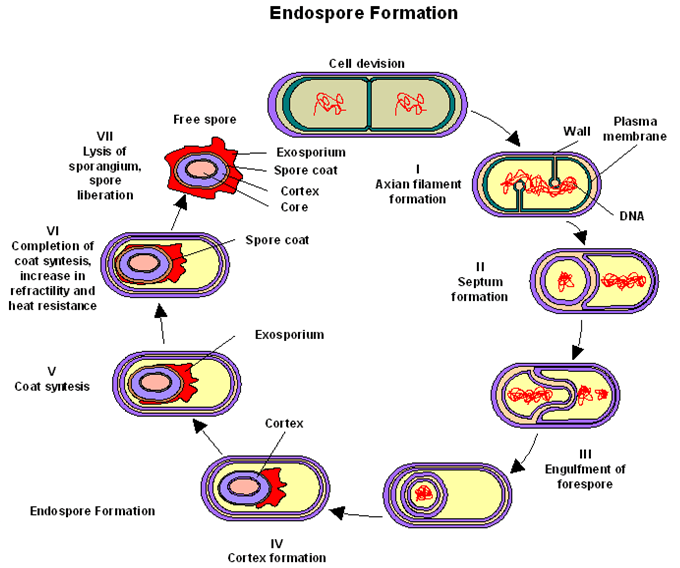
Paso de espora a célula vegetativa:
Activación: proceso reversible, preparación de la espora para la germinación, se da por algún estímulo, por ejemplo el calor.
Germinación: es un proceso rápido, se hincha, la cubierta se rompe o se absorbe. Se pierde la resistencia al calor, radiación, desecación, etc. Se desencadena por ciertos metabolitos, por ejemplo los nutrientes del medio (aminoácidos, azúcares, etc.).
Crecimiento: protoplasto fabrica nuevos componentes.
Estructuras externas:
Pared celular:
Las bacterias, a excepción de algunos micoplasmas y algunas arquibacterias, tienen una pares fuerte que les da forma y las protege de la lisis osmótica y sustancias tóxicas. En algunos microorganismos puede tener componentes que le confieren patogenicidad.
En 1884 el investigador danés Christian Gram observó diferencias tintoriales relacionadas con la habilidad de ciertas células bacterianas de retener el colorante cristal violeta después de la decoloración con etanol al 95%, estas células fueron llamadas Gram positivas. Las células que se decoloraban y solamente son visibles al teñirlas con un colorante de contraste son referidas como Gram negativas.
Esto se debía a las diferencias existentes en la estructura de la pared celular:
- Gram positiva: formada por una única capa homogénea de 20-80 nm de grosor, de peptidoglucano o mureína situado por fuera de la membrana plasmática. Además de polímeros aniónicos (ácidos teicoicos y teicourónicos), polisacáridos neutros, proteínas y en algunos casos lípidos. Los ácidos teicoicos se encuentran presentes en la pared de dos maneras, unidas covalentemente al peptidoglucano o a lípidos de membrana como ácidos lipoteicoicos. Sus funciones son unir la pared celular con la membrana citoplasmática y acumular cargan negativas para atrapar cationes como el Mg2+, que es esencial para la actividad normal de las enzimas de membrana. Existe un espacio periplásmico que no tiene tantas proteínas como en las Gram negativas, pero secretan exoenzimas.
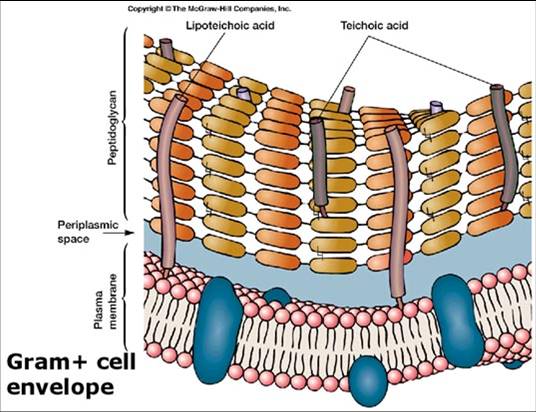
- Gram negativa: es más compleja, posee una capa de 2-7 nm de grosor de peptidoglucano rodeada por una membrana externa de unos 7-8 nm. El espacio entre estas dos capas se denomina espacio periplásmico y la sustancia que ocupa ese lugar periplasma. En este espacio hay muchas proteínas que participan en la captación de nutrientes y proteínas quelantes que participan en el transporte de materiales hacia el interior de la célula, contiene también enzimas que participan en la síntesis del peptidoglucano y en la modificación de compuestos tóxicos que podrían lesionar a la célula.
Por fuera de la fina capa de peptidoglucano se encuentra una membrana externa que cumple la función de una membrana lisosomal, las enzimas hidrolíticas del espacio periplásmico se mantienen separadas de la célula gracias a la membrana y al peptidoglucano para prevenir la autodigestión.
El espacio periplásmico es isoosmótico con el citoplasma y fuertemente aniónico con el medio externo, por lo que es en parte responsable de mantener la osmolaridad (afuera es hiposomótico). La membrana externa funciona como una barrera permeable para los solutos hidrofilicos.
El componente más importante de la membrana externa es el Lipopolisacárido (LPS) o endotoxina que es la principal toxina de las bacterias entéricas patógenas. Los fosfolípidos y proteínas se encuentran en menor concentración que en la membrana plasmática. Existe un tipo de proteínas llamadas porinas que forman canales transmembrana y permiten el paso de solutos de acuerdo al peso molecular (nutrientes hidrofílicos).
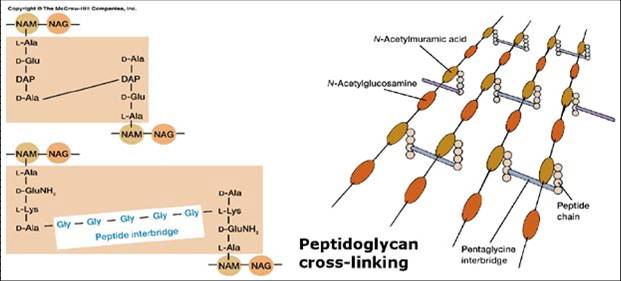
Peptidoglucano: Polímero constituido por residuos alternantes de N-acetilglucosamina y ácido N- acetilmurámico (aminoazúcares). Una cadena de 4 aminoácidos D- y L- alternantes está conectada al ácido N- acetilmurámico y varía de acuerdo a las especies.
Glicocálix:
Polisacáridos de origen bacteriano que se localizan alrededor de la membrana externa (Gram negativas) y del peptidoglucano Gram positivas).
Se clasifican como:
- Slime o tipo S: matriz difusa
- Cápsula: matriz fibrosa bien organizada:
rígida
flexible
integral
periférica
La cápsula está formada por polisacáridos y en algunos casos puede contener proteínas, su componente principal es el agua lo que puede proteger a la bacteria de la desecación. Permite a las bacterias unirse a diferentes tipos de sustratos, previene la ingestión de la bacteria por las células fagocíticas, evita virus bacterianos y puede dar patogenicidad.

Pilis y fimbrias:
En la mayoría de bacterias Gram negativas se encuentran apéndices cortos, finos y más delgados que un flagelo que no están involucrados en la movilidad:
- Fimbrias: Tubos delgados compuestos de subunidades proteicas ordenadas de forma helicoidal, diámetro de 3-10 nm y unos μm de largo. Al menos 1 000 fimbrias, son únicamente visibles en un microscopio electrónico debido a su tamaño. No se conoce con certeza su función, se ha observado que estas estructuras pueden servir para anclar a la bacteria a superficies sólidas como rocas y tejidos.

- Pili: apéndice similar que se encuentra en una menor cantidad en la célula (de 1-10 por célula) y es de mayor tamaño (9-10 nm de diámetro). Están genéticamente determinados por factores sexuales o plásmidos conjugativos y son importantes para el apareamiento bacteriano.

Flagelos:
La movilidad: requiere un gasto de energía y dentro de un hábitat tiene importantes consecuencias ecológicas y diferencia entre la vida o la muerte de la célula. El movimiento a lo largo de superficies sólidas por deslizamiento o por vesículas de gas en medios acuáticos, no es un sistema de movilidad verdadera. La capacidad de movimiento independiente está dada por el flagelo.
Es un apéndice locomotor delgado y semirígido de 20 nm de diámetro y de 15-20 μm de largo.
Son tan delgados que no son visibles al microscopio de luz, pero pueden utilizarse tinciones especiales para así aumentar su diámetro. Su estructura detallada puede ser únicamente visible con un microscopio electrónico.
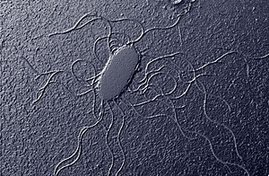
Existen diversos patrones de distribución flagelar, que son muy útiles para la identificación bacteriana:
- Monotrico: un solo flagelo colocado al final de la célula bacteriana y se denomina flagelo polar
- Anfitrico: un flagelo en ada polo celular

- Lofotrico: un grupo de flagelos en uno o varios polos celulares

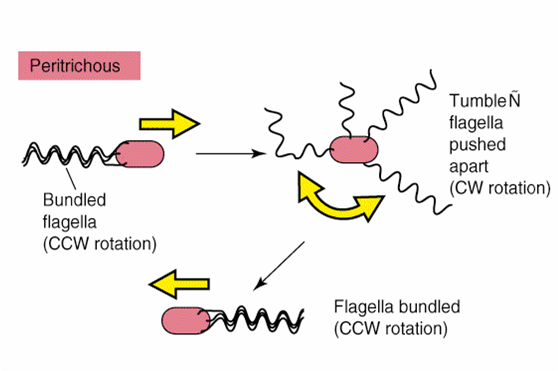
- Peritrico: Falgelos por toda la superficie bacteriana

Utraestructura flagelar:
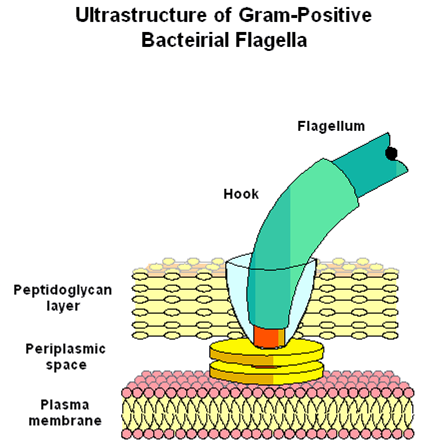
El flagelo está compuesto por 3 partes fundamentales:
-Filamento: porción más larga, cilindro rígido y hueco constituido de una única proteína llamada Flagelina.
-Gancho: une el filamento con el cuerpo basal como una cubierta flexible, ligeramente más ensanchado, constituido de subunidades proteicas diferentes.
-Cuerpo Basal: embebido en la célula, es la parte más compleja del flagelo:
a.Gram negativas: cuerpo basal con 4 anillos conectados a un filamento central. Los anillos más externos L y P se asocian con la capa de lipopolisacáridos y peptidoglucano, respectivamente y los internos S y M con la membrana plasmática.
b.Gram positivas: cuerpo basal con únicamente 2 anillos, el más interno asociado a la membrana plasmática y el externo al peptidoglucano.
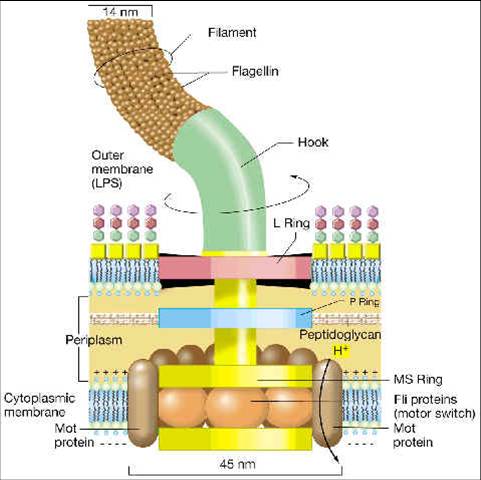
Síntesis Flagelar:
Proceso muy complejo que involucra varios genes, al menos unos 20-30 genes: Genes que codifican para la flagelina, 10 o más genes que codifican para las proteínas del gancho y el cuerpo basal
Otros genes están involucrados con el control de la construcción o la función flagelar.
Aún no se sabe cómo la célula regula o determina la localización exacta del flagelo. El crecimiento del flagelo no se produce a partir de la base sino de la punta
Las moléculas de flagelina formadas en la célula pasan a través de un núcleo hueco del flagelo y se insertan en el extremo terminal, luego las subunidades espontáneamente se agregan. El crecimiento del flagelo se produce más o menos continuamente, aunque la velocidad de crecimiento disminuye a medida que aumenta el tamaño del filamento.
Si se rompe una parte de la punta, se regenera.
Mecanismo de Movimiento Flagelar:
Mecanismo de Movimiento Flagelar:
Cada flagelo es una estructura semirrígida incapaz de flexionarse pero, se mueve por rotación que parte del cuerpo basal que funciona como un motor. La energía que se necesita para la rotación del flagelo proviene de la fuerza motriz generada por un gradiente de protones.
Los flagelos no rotan a velocidad constante sino que la velocidad de rotación aumenta o disminuye en relación con la intensidad de la fuerza motriz del gradiente. La dirección de la rotación flagelar determina la naturaleza del movimiento bacteriano.
-Monotrico: el flagelo polar rota en sentido contrario a las manecillas del reloj para un movimiento hacia adelante. Para detenerse o dar vuelta cambia la rotación flagelar en sentido de las manecillas del reloj.
-Monotrico: el flagelo polar rota en sentido contrario a las manecillas del reloj para un movimiento hacia adelante. Para detenerse o dar vuelta cambia la rotación flagelar en sentido de las manecillas del reloj.

-Peritrico: la rotación es parecida.

1 comentario:
Casino at the Hard Rock Resort & Casino
Welcome to the Hard 성남 출장안마 Rock 동두천 출장마사지 Hotel & Casino, with over 고양 출장샵 3,500 slot machines, 70 table games and 밀양 출장안마 the newest addition is 전라남도 출장안마 Hotel of the Year!
Publicar un comentario